
Sahelanthropus è il più antico genere di ominino appartenente al lignaggio umano, scoperto da Brunet nel 2001. Attraverso metodi di datazione biocronologici, quest’ominino risulta essere antico di circa 7-6 milioni di anni. A questo genere è associata solo una specie, Sahelanthropus tchadensis, un taxon di importanza fondamentale rinvenuto in Chad, nel sito di Toros-Menalla nell’Africa centro-occidentale. Quest'area, situata subito a sud del Sahara, in epoche antiche era caratterizzata da un habitat complesso, con fiumi costeggiati da foreste, boscaglie erbose e una serie di laghi. I sedimenti depositati lungo queste antiche spiagge o nei laghi hanno preservato una ricca varietà di organismi, tra cui pesci d’acqua dolce e erbivori tipici della boscaglia o delle foreste, testimonianza della ricchezza di questo ambiente. È importante sottolineare che di questa specie è stato trovato un cranio, sebbene deformato. Grazie alla paleontologia, è possibile correggere queste deformazioni di origine tafonomica. Sono stati rinvenuti anche un paio di mandibole che forniscono informazioni cruciali. Ad esempio, il cranio indica che questa specie possedeva un cervello simile a quello di uno scimpanzé. La parte superiore della faccia e i rilievi sopraorbitali sono tipici degli ominini, mentre la mandibola risulta essere più spessa di quella di uno scimpanzé moderno. Inoltre, è interessante notare che i canini sono consumati solo nella parte superiore, a differenza degli scimpanzé, i cui canini presentano consumo anche lateralmente. Esistono ancora scetticismi, principalmente dovuti all'esiguo numero di fossili, che non consente di delineare un identikit soddisfacente di questa specie. In passato, alcuni ritenevano potesse trattarsi di un antico gorilla, o addirittura dell’antenato comune dello scimpanzé e dell’uomo. Tuttavia, alcune ricerche recenti concordano sul fatto che si trattasse di un ominino appartenente al lignaggio umano. Al momento, S. tchadensis è considerato uno dei primi ominini rinvenuti in Africa centro-occidentale, suggerendo che essi occupassero un’area più ampia di quanto precedentemente ipotizzato.
 |
| Cranio di Sahelanthropus tchadensis Didier Descouens, CC B-SA 4.0 (Wikimedia Commons) |
 |
| Cranio di Sahelanthropus tchadensis. (A) visione frontale, (B) visione laterale, (C) visione dorsale, (D) visione basale (Brunet et al., 2002) |
 |
| Breve elenco di alcune caratteristiche primitive e derivate di Sahelanthropus tchadensis |
In generale, le dimensioni del cervello, la postura eretta e l’andatura bipede (seppur con diverse sfaccettature) indicano se ci troviamo davanti a un ominino. Altri caratteri, seppur diversificati, vengono presi in considerazione, delineando un insieme di caratteristiche arcaiche e derivate acquisite dal lignaggio umano nel corso del tempo, configurando così un’evoluzione a mosaico. In parole povere, alcuni di questi tratti antichi possono anche comparire in individui più recenti, specialmente nelle cosiddette “specie insulari”, insieme a tratti appena sviluppati. Questo sottolinea che l’evoluzione non segue un percorso lineare; nel corso del tempo, alcune caratteristiche possono scomparire, mentre altre possono emergere e stabilirsi all’interno delle popolazioni, oppure scomparire e riapparire in generazioni successive. Esistono prove, sia dirette che indirette, che suggeriscono che Sahelanthropus fosse bipede. Ad esempio, la presenza di un foro occipitale in posizione avanzata è associata a una locomozione bipede, poiché il cranio è posizionato su una colonna vertebrale relativamente perpendicolare al terreno. Inoltre, come accennato in precedenza, le caratteristiche dei denti indicano che non si tratta né di uno scimpanzé né di un gorilla, ma di un ominino. Questo è evidenziato dalle ridotte dimensioni del canino e da uno smalto dei molari più spesso, rispetto a quelli di Gorilla e Pan. Tuttavia, c'è incertezza sul fatto che Sahelanthropus fosse un ominino o un ominide. Ad esempio, nello stesso sito è stato ritrovato un femore, descritto nel 2020, la cui morfologia sembra non corrispondere a quella di un ominino con una locomozione bipede abituale, sollevando dubbi sulla sua associazione con Sahelanthropus. In breve, il dibattito è in parte aperto, ma la presenza di un'andatura bipede non necessariamente rappresenta un confine netto.
Una recente ricerca ha cercato di gettare luce sul periodo temporale in cui vissero S. tchadensis e Australopithecus bahrelghazali, un ominino che compare successivamente e che abitò gli stessi luoghi di Sahelanthropus. Questo aspetto è particolarmente interessante data la posizione geografica del Ciad. Attraverso la datazione con nuclidi cosmogenici di queste specie, è stato possibile, in primo luogo, datare i luoghi di rinvenimento delle stesse (deserto di Djurab nel Ciad settentrionale). Questa datazione è stata agevolata anche grazie ai fossili di mammiferi presenti, i quali hanno permesso di stimare la (bio)cronologia di questi resti di ominini. La traccia di 10 Be/ 9 Be rinvenuta in una pelite indica che lo strato in cui è stata scoperta A. bahrelghazali ha un'età di 3,58 ± 0,27 milioni di anni, suggerendo che questa specie fosse contemporanea dell'Australopithecus afarensis (Lucy). Per quanto riguarda il sito in cui è stata ritrovata S. tchadensis, sempre mediante lo stesso metodo di studio basato su 10 Be/ 9 Be, le analisi condotte sull'unità di antracoteridi contenente la staffa dell'ominino indicano che lo strato possiede un'età compresa tra 6,8 e 7,2 milioni di anni.
In generale, i dati dimostrano innanzitutto l'accuratezza dell'età autigenica ottenuta tramite il rapporto 10 Be/9 Be, confermata dal confronto con le stime biocronologiche basate sull'evoluzione dei lignaggi fossili dei mammiferi. Questo dimostra che l'ambiente ha svolto un ruolo fondamentale nella conservazione di questi nuclidi cosmogenici. L'ambiente circostante era caratterizzato da periodi aridi intervallati da periodi umidi, durante i quali si formò un grande lago poco profondo. Questo lago ha fornito sedimenti olocenici che hanno permesso di determinare la quantità iniziale di 10 Be/9 Be, preservando i sedimenti più antichi in una sorta di sistema chiuso, simile a un termos. Inoltre, la presenza di peliti autigeniche, datate anch'esse tramite il rapporto 10 Be/9 Be e conservate nell'arenaria argillosa contenente la mandibola di A. bahrelghazali, indica che la località di Toro Koro ha almeno 3,3 milioni di anni. Questo non solo permette di calibrare gli strumenti utilizzati per la datazione, ma fornisce anche informazioni rilevanti per l'orologio molecolare. Indica infatti che S. tchadensis è sicuramente un ominino più giovane di 8 milioni di anni e che la divergenza tra i lignaggi di Pan e Homo risale a almeno 8 milioni di anni fa. Ciò è supportato anche dal ritrovamento di Chororapithecus abyssinicus, un "paleogorillidae" antico di 10 milioni di anni (Suwa et al., 2007). Il mosaico di caratteri plesiomorfi e apomorfi di S. tchadensis è probabilmente molto vicino al tempo di divergenza tra Pan e Homo, e non supporta l'idea che questi lignaggi si siano separati attorno ai 6,3 milioni di anni, né che la presenza di un mosaico di morfologie sia dovuta a un'ibridazione tra individui appena separati di Pan e Homo, prima di separarsi definitivamente (Patterson et al., 2006).
 |
| Posizione geografica del Chad |
La fauna dell'antico Chad è estremamente variegata e comprende una vasta gamma di habitat, inclusi quelli lacustri. Le fonti d'acqua presenti ospitano diverse specie di pesci. Tra queste, la famiglia Latidae (Perciformes), attualmente presente in Australia, Giappone e nei grandi laghi africani, è particolarmente affascinante. Questi pesci sono noti per la loro capacità di sopravvivere in ambienti sia salmastri che dolci e marini, ma nel caso del ritrovamento in Chad, è probabile che gli individui siano rimasti intrappolati a seguito di abbassamenti più o meno rapidi del livello dell'acqua, necessitando di acque ricche di ossigeno. Un'altra famiglia di pesci presente è la Ariidae (Siluriformes), adattata a condizioni prevalentemente anossiche. È interessante notare anche la presenza di Hydrocynus (pesce tigre), un predatore che caccia a vista nelle acque profonde e ben ossigenate. La diversità di specie e adattamenti evidenzia la ricchezza e la complessità degli habitat acquatici presenti nell'antico Chad.
.png) |
| Mappa del sito fossilifero di Toros-Menalla. Modificata da Vignaud et al., (2002) |
La presenza di legno silicizzato con un grande diametro suggerisce una somiglianza con le liane attuali, indicando la presenza di gallerie di foreste lungo i laghi. Questi habitat erano popolati da una vasta varietà di animali, tra cui Python sebae, noto anche come pitone delle rocce africane. Questo grosso animale, che può raggiungere una lunghezza media di 3-4 metri, era in grado di muoversi sia tra le piante vicino alle fonti d'acqua che di nuotare, potendo rimanere immerso anche per lungo tempo. È interessante notare che si nutriva anche di coccodrilli. Crocodylia mostra una grande biodiversità, con Codrocylus lloydii cche risulta essere il più rappresentativo, mentre Gavialidae era caratterizzata dalla presenza di predatori generalisti. Inoltre, c'erano specie come il semi-acquatico Varanus niloticus, tartarughe, lucertole, che caratterizzavano la parte superiore della sezione di Toros-Menalla. Questi ambienti erano abitati da grandi animali che raggiungevano almeno il metro di lunghezza, principalmente pescivori, indicando la presenza di bacini idrici estesi e prominenti, con aree paludose e ben vegetate. La presenza del pesce Polypterus, che tollera ambienti poveri di ossigeno, e di Gymnarchus, che vive in acque paludose e torbide, dove un sistema sensoriale elettrico è vantaggioso, suggerisce ulteriormente la natura variegata e ricca di questi ambienti. Infine, non dobbiamo dimenticare che queste "coste" erano anche abitate da grandi ippopotami, completando il quadro di una fauna diversificata e adattata agli habitat acquatici e paludosi dell'antico Chad.
Per quanto riguarda i mammiferi, l'elenco è davvero lungo. Sono presenti esemplari di Lagomorpha, di dimensioni più piccole rispetto a quelli rinvenuti a Laetoli, oltre a equidi simili ad Hipparion, giraffidi, proboscidati e carnivori (in gran parte iene di grandi dimensioni), soprattutto iene di varie dimensioni, mustelidi(in particolar modo la lontra). Per quanto riguarda i primati, escludendo Sahelanthropus, vi è la presenza anche di alcune colobine non ben definite. Di particolare interesse è la scoperta del primo scheletro del Neogene africano associato a un mammifero tubulidentato, un insettivoro caratterizzato anche dalla presenza di termitai. L'unità stratigrafica in cui è stato rinvenuto Sahelanthropus è informalmente chiamata "unità degli antracoteridi", a causa dell'abbondanza di fossili associati a questa famiglia di artiodattili imparentati sia con gli ippopotamidi sia con i suidi, in particolare riferiti a Libycosaurus petrocchii. I bovidi rappresentano circa il 55% dei resti di mammiferi della località, mentre i mammiferi anfibi costituiscono circa il 28%. I denti a corona relativamente alta di tutti i bovidi e l'assenza di tragelaphine e boselaphine (anch'essi bovidi) sono chiare evidenze della presenza di praterie aperte. Questo insieme faunistico del sito degli ominini è compatibile con una diversità di habitat, tra cui praterie aperte (bovidi), savane boschive (proboscidati e giraffidi), ambienti di acqua dolce (pesci, tartarughe, serpenti, coccodrilli, antracoteridi, ippopotami e lontre) e probabilmente foreste a galleria (primati). Gli ominini più antichi come Orrorin e Ardipithecus sono contemporanei a faune legate ad ambienti boscosi, mentre le prime australopitecine hanno vissuto in una vasta gamma di habitat. Al contrario, la fauna di Toros-Menalla suggerisce la presenza di un ambiente a mosaico che va dalla foresta a galleria ai margini dei bacini lacustri fino alla presenza di un grande ambiente di savana e prateria. Determinare l'habitat preciso in cui visse Sahelanthropus è difficile, e la scarsità di informazioni sulla locomozione non aiuta. È possibile che abbia vissuto vicino a un lago, non lontano da un deserto sabbioso. La parte centrale della sezione corrisponde a un ambiente perilacustre poco profondo, soggetto a frequenti inondazioni. In questa fascia vegetata sono presenti numerosi ambienti vicino a un deserto sabbioso, ma le tracce di paleocorrenti che si sviluppano in tutte le direzioni indicano la presenza di un sistema tortuoso e complesso. I sedimenti del bacino suggeriscono che al tempo non fossero presenti sistemi fluviali perenni.
Le più antiche tracce di bipedismo?
La questione del bipedismo è sempre stata oggetto di dibattito poiché inizialmente si riteneva essere una caratteristica esclusiva del genere Homo. Tuttavia, è emerso che il bipedismo è stato un tratto significativo per numerosi ominini, incluse le australopitecine, così come per il nostro "parente molto alla lontana" Oreopithecus bambolii. Questo non è stato un fenomeno graduale e il bipedismo era già presente in un ominino che tradizionalmente era considerato principalmente arboricolo, ovvero Sahelanthropus. Le prove di ciò provengono sempre dal Ciad (Toros-Ménalla) e sono datate circa 7,2-6,8 milioni di anni fa (Miocene). Queste prove derivano dallo studio di componenti postcraniali come il femore e l'ulna (Daver, 2022), indicando che questo ominino era in grado sia di camminare in posizione eretta che di arrampicarsi sugli alberi, utilizzando la locomozione più adatta a seconda del contesto.
Il femore di S. tchadensis indica che questo ominino era caratterizzato da una locomozione bipede regolare. Questo è evidente dai contorni della sezione trasversale del femore, dalla distribuzione dell'osso corticale lungo e dalla struttura femorale relativamente attorcigliata. Queste caratteristiche favorirebbero la dissipazione dei carichi di compressione causati dal bipedismo. Come per altri ominini, vi era una protolinea aspera ben definita, che aumenta la robustezza dell'osso e fornisce un punto solido di attacco per i muscoli. La presenza della tuberosità glutea laterale e della platimeria sottotrocanterica, senza la presenza della fossa ipotrocanterica o inferolaterale, sono in genere caratteristiche associate a una maggiore flessione-estensione dell'anca. Queste caratteristiche sono tipiche di un ominino arcaico, ma funzionalmente indicano comunque una capacità di locomozione bipede abituale su substrati terrestri. Ora prendiamo in considerazione l'ulna, sempre proveniente dal Ciad, che mostra una serie di caratteristiche morfologiche associabili a un comportamento perlopiù arboreo, simile a quello di A. ramidus, di O. tugenensis e di Ardipithecus kadabba. È importante notare, tuttavia, che lo stile di vita arboreo di molti primati non è uniforme in termini di locomozione. Infatti, l'ulna di S. tchadensis indica che questa specie è stata caratterizzata da un arrampicamento di tipo pronogrado e ortogrado, che comportava comunque una presa sicura degli arti. In generale, non si osservano nell'osso attività sospensive abituali come l'oscillazione e/o la sospensione dell'avambraccio. Quindi, per riassumere, i dati del femore suggeriscono che l'arrampicata ortograda nel contesto arboreo facesse parte del repertorio di comportamento di questa specie, e che svolgesse una funzione di "carico" per gli arti posteriori (come descritto per A. ramidus e, probabilmente, O. tugenensis). Quindi, Sahelanthropus conserva le prime prove di bipedismo abituale nel lignaggio umano (ominini), confermando i precedenti dati ottenuti dal materiale cranico basati sull'orientamento del piano orbitale, del forame magno e sulla morfologia del piano nucale. Il bipedismo è una forma di locomozione che caratterizza anche primati non-ominini, come O. bambolii; tuttavia, in questo caso possiamo dire che questo adattamento su substrati terrestri possa essere comparso poco tempo dopo la divergenza tra i lignaggi dell'uomo e dello scimpanzé, probabilmente in concomitanza con adattamenti arborei. Anche le australopitecine erano in grado, in generale, di scegliere la tipologia di locomozione (bipede o arborea) in base al contesto ambientale, e anche le prime specie umane come Homo habilis e Homo rudolfensis non sono da meno. Senza contare che anche specie più recenti o derivate come Homo naledi erano in grado di adattarsi e scegliere la tipologia di locomozione in base alle circostanze ambientali.
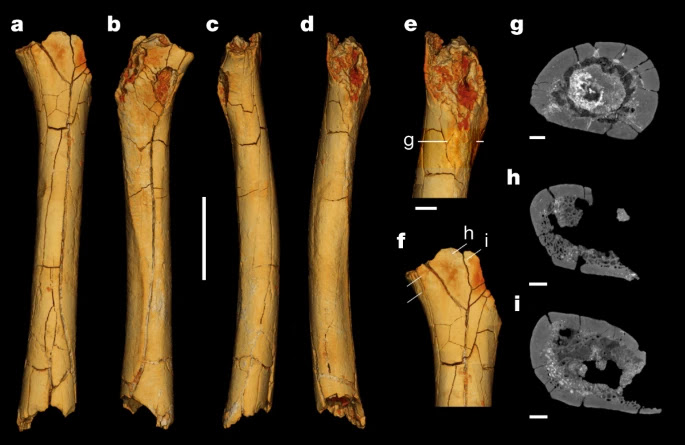 |
| Femore di Sahelanthropus tchadensis rappresentato virtualmente (Daver et al., 2022) |
 |
| Femori di Sahelanthropus tchadensis, uomo moderno, scimpanzé. scimpanzé e orango. (A) visione anteriore, (B) visione posteriore, (C) visione laterale e (D) posteriore (Daver et al., 2022) |
 |
| Ulna di Sahelanthropus tchadensis (Daver et al., 2022) |
Va fatta una piccola considerazione anche sugli altri ominini. Orrorin è associato ad aree boschive aperte con significativa copertura arborea; Ar. kadabba (5,2 milioni di anni circa) è associata a un ambiente caratterizzato sia dalla presenza di boschi che di praterie umide; Ar. ramidus (4,4 milioni di anni circa) abitava, molto probabilmente, in un bosco erboso alimentato da acque sotterranee (probabilmente palmeti). I primi ominini dell'Africa orientale vivevano generalmente in un contesto arboreo, ma il sito di Toros-Ménalla era caratterizzato da foreste chiuse, palmeti e ambienti erbosi misti, condividendo una componente arborea nel loro ambiente. Per quanto riguarda l'area fossilifera, i dati suggeriscono un paesaggio eterogeneo che comprende foreste chiuse (probabilmente foreste ripariali), palmeti e ambienti erbosi misti. Si poteva passare dai boschi alle savane e praterie acquatiche in pochi chilometri. Quindi, una specie come S. tchadensis (e anche gli altri ominini del Ciad) era in grado di sfruttare sia ambienti arborei che terrestri, sia per procurarsi il cibo che per accedere a risorse idriche. Un repertorio locomotore così diversificato, e la presenza di habitat boschivi in un range temporale che abbraccia circa 2,5 milioni di anni, suggerisce che la nicchia ecologica di questi primi ominini non era necessariamente legata all'espansione delle aree aperte e relativamente secche; pertanto, si tratterebbe di una specie opportunistica che dipendeva sia dalle risorse terrestri che arboree.
Possiamo concluderla qui? Il lavoro di cui ho appena parlato è stato oggetto di intense critiche a causa della presunta inconsistenza dei dati, con molti ricercatori che mostrano quanto siano più consistenti i dati del 2020. Vediamo cosa dicono. Si tratta di un femore parziale sinistro e, da un'analisi tafonomica, sembra che la diafisi femorale possa appartenere a una specie di primate sconosciuta, in generale a un "ominide" che non mostrava abitudini bipedi. Tuttavia, in base al principio di parsimonia, potrebbe essere ragionevole supporre che appartenga a S. tchadensis, soprattutto considerando un'analisi comparativa con il femore di O. tugenensis, datato al tardo Miocene e proveniente dal Kenya, che suggerisce una specie distinta. Potrebbe essere plausibile considerare S. tchadensis come un "ominide" in senso lato, un possibile "antenato" degli ominini che non ha attualmente alcun rappresentante vivente. Tuttavia, analisi più approfondite potrebbero portare a una maggiore comprensione di questa specie e determinare se sia un ominino in senso stretto. Dato che il lignaggio umano presenta una vasta diversità tassonomica, è difficile stabilire con certezza le sue relazioni filogenetiche. È possibile che ci siano stati fenomeni evolutivi, come la deriva genetica, che hanno portato i discendenti di un antico ominino bipede a sviluppare una locomozione bipede non abituale. In definitiva, S. tchadensis rimane in una sorta di limbo per alcuni ricercatori, ma si spera che ulteriori ricerche possano fornire maggiori chiarimenti sulla sua posizione filogenetica e sul suo ruolo nell'evoluzione umana, che potrebbe essere un ominino in senso stretto, un antico parente degli scimpanzé o persino un antenato comune del lignaggio Pan-Homo.
.png) |
| Femore di Sahelanthropus tchadensis (Macchiarelli et al., 2020) |
Ora, passiamo agli arti superiori e al cranio, perché, volente o nolente, sono stati forniti un sacco di dettagli interessanti che rendono il cranio di Sahelanthropus, se considerato un ominino, il più antico associato al lignaggio umano. Questo ha permesso ai ricercatori di fare una prima analisi "parentale" con gli ominini successivi, i gorilla e gli scimpanzé. Quanto complicano le cose! I risultati indicano innanzitutto che Sahelanthropus è sicuramente un "ominide" che condivide molte caratteristiche primitive con gli scimpanzé, ma nel complesso è molto simile all'Australopithecus, soprattutto per quanto riguarda il basicranio. Inoltre, Sahelanthropus si distingue dalle australopitecine per la combinazione di una breve regione sottosfenoidea associata a una faccia superiore verticale che sporge davanti al neurocranio. È stato un lavoro in parte difficile perché il campione mostra una parziale distorsione dovuta a fratture e deformazioni plastiche, ma grazie alla paleontologia virtuale, i problemi sono stati risolti. Questo ha confermato che Sahelanthropus è una specie caratterizzata da un mosaico di morfologie, sia derivate (come nelle australopitecine) che primitive, e che probabilmente era bipede (anche se non in modo obbligato o abituale). Questo ha permesso di stabilire un collegamento tra Sahelanthropus e gli altri ominini menzionati precedentemente, ma il risultato suggerisce che potrebbe non essere direttamente imparentato con loro. Vediamo ora alcune di queste caratteristiche morfologiche:
La lunghezza, la proiezione del viso, la posizione anteroposteriore relativa del forame magno e delle fosse mandibolari, e la lunghezza e l'altezza della volta cranica avvicinano Sahelanthropus alle australopitecine.
- Rispetto a Pan e Gorilla, Sahelanthropus presenta una faccia più verticale con un rostro antero-posteriore relativamente più corto e una base cranica posteriore in una posizione più anteriore.
- Il contorno mediosagittale del neurocranio e l’altezza relativa della faccia superiore separano Sahelanthropus da A. afarensis, Paranthropus boisei e da Australopithecus africanus.
- La forma della volta cranica assomiglia a Pan troglodytes e Gorilla gorilla per via del contorno superiore basso e lungo del neurocranio, soprattutto nella parte anteriore all'apice della volta.
- Il piano nucale di Sahelanthropus, tuttavia, è relativamente lungo, piatto e ruotato verso l'orizzontale come in A. africanus e A. afarensis.
- L'altezza facciale relativa (dalla nasion alla prosthion) in Sahelanthropus è simile a quella di A. africanus e A. afarensis e H. habilis, ma più corta di quella di G. gorilla e P. troglodytes.
- Sahelanthropus presenta alcune differenze, forse derivate, dai crani di Australopithecus e delle scimmie antropomorfe africane, in particolare nella faccia superiore (comprese le sovrastrutture orbitali, la glabella e il nasion), che sporge sostanzialmente davanti al neurocranio. Inoltre, la faccia di Sahelanthropus ha una premascella relativamente corta senza diastema e un prognatismo subnasale ridotto rispetto non solo a Pan e Gorilla ma anche ad alcuni rappresentanti cranici dell'Australopithecus.
- Gli aspetti della struttura complessiva del viso di Sahelanthropus, ovvero faccia superiore sporgente e prognatismo sottonasale ridotto, assomigliano a quelli di H. habilis.
- Sahelanthropus condivide molte caratteristiche apparentemente primitive con Pan e Gorilla, in particolare nel palato e nelle regioni della volta cranica diverse dal piano nucale. Ha anche un volume endocranico stimato di 360–370 cc, il più piccolo finora documentato per un ominino adulto, ma compreso nell'intervallo degli scimpanzé.
- Sahelanthropus mostra diverse sinapomorfie da ominino tra cui un piano nucale lungo e piatto che è più orientato orizzontalmente rispetto alle scimmie africane; un basioccipitale accorciato; e un forame magno posizionato più anteriormente. Inoltre, la faccia di Sahelanthropus ha una premascella relativamente corta senza diastema e un prognatismo subnasale ridotto rispetto non solo a Pan e Gorilla ma anche ad alcuni rappresentanti cranici dell'Australopithecus.
- Sahelanthropus condivide anche diverse caratteristiche craniche derivate con gli ominini successivi, specialmente nella volta cranica posteriore e nella base cranica, diverse da qualsiasi Australopithecus conosciuto. Molte di queste caratteristiche, come l’altezza sottonasale, la protrusione palatale e la relativa proiezione facciale superiore, possono essere derivati in modo diverso rispetto a quelli dell'Australopithecus. Altri sono probabilmente primitivi o convergenti con Homo. Ad esempio, le porzioni superiori e mediane del volto di Sahelanthropus sono simili in larghezza, come in Pan e Homo, ma diversamente dalla configurazione a forma esagonale dell'Australopithecus, in cui la parte mediana del volto è più larga di quella superiore.
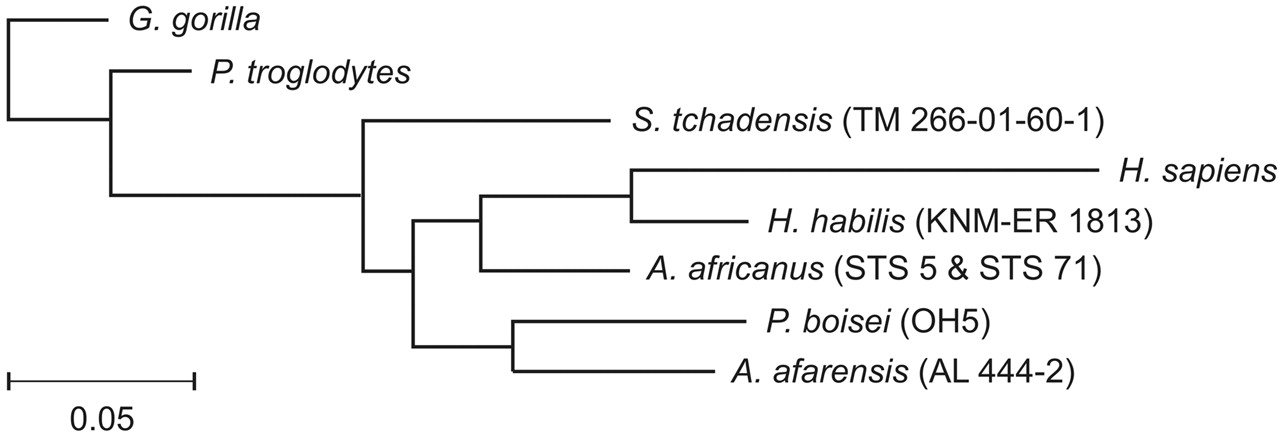 |
| Analisi fenetica delle affinità tra Sahelanthropus tchadensis e Pan, Gorilla e gli ominini (Guy et al., 2005) |
.gif) |
| Questo è un grafico scritto in inglese nel quale sono state aggiunte altre possibili specie, non ancora riconosciute come tali. Differisce da quello che si trova in genere sui libri del Paleoantropologo Giorgio Manzi, ma comunque mostra le varie affinità morfologiche delle varie specie e il susseguirsi di esse nel corso del tempo. Essendo la nostra specie l'unica del genere Homo in vita, viene utilizzata come metro di comparazione e, quando una specie presenta delle affinità, si troverà più a sinistra; se mostra caratteristiche differenti e divergenti, la specie tenderà ad essere posizionata verso destra. In questo caso, Sahelanthropus tchadensis si trova in una posizione particolare in quanto è molto "lontano" dalle morfologie tipiche di Homo, se non per alcune caratteristiche, ma nel complesso non è una specie distante quanto P. boisei. (Wood et al., 2020) |
La forma cranica tridimensionale suggerisce che S. tchadensis si raggruppi feneticamente con gli ominini successivi piuttosto che con Pan, ma nel complesso non è più simile a qualsiasi altra specie di ominini più derivata. Si presume, quindi, che faccia parte di un clade di ominini primitivi ancestrali di tutti i generi successivi, inclusi Australopithecus e Homo. Altri membri di questo clade includono potenzialmente Ardipithecus e Orrorin. Inoltre, vi è anche il problema della paleobiogeografia degli ominini del Miocene superiore. Pertanto, sebbene Ardipithecus e Orrorin siano entrambi conosciuti nell'Africa orientale (rispettivamente in Etiopia e in Kenya), Sahelanthropus è stato rinvenuto nell'Africa centrale (Ciad), a circa 2.500 km a ovest della Rift Valley. Questo modello solleva interrogativi interessanti sulla filogenesi degli ominini. Infatti, tra Libia e Ciad ci sono stati scambi faunistici 7 milioni di anni fa, come attestano alcuni mammiferi, ma non sembra ci sia traccia di scambi faunistici tra il Ciad e l’Africa Orientale. Tutti quegli interrogativi del tipo “è un ominino o un ominide, oppure uno scimpanzé” servono proprio a far capire che, nonostante una moltitudine di dati, non è facile collocare una data specie in un determinato gruppo. Chi ha svolto questo lavoro, per esempio, indica che si tratta chiaramente di un ominino caratterizzato da un complesso mosaico di caratteristiche, a differenza del lavoro del 2020 che colloca Sahelanthropus in una sorta di limbo. A questo studio, possiamo aggiungerne un altro che riguarda la ricostruzione virtuale del cranio quasi completo, dei denti e di 3 frammenti mandibolari di S. tchadensis. Il cranio è interessante dal punto di vista della sistematica e ciò che ne consegue è che questa specie è a tutti gli effetti un ominino e non è più strettamente imparentato con le grandi scimmie africane. L'analisi del basicranio, inoltre, indica che S. tchadensis potrebbe essere stato un bipede eretto, suggerendo che il bipedismo era presente nei primi ominini conosciuti e probabilmente sorse subito dopo la divergenza tra i lignaggi degli scimpanzé e degli umani. Anche se ciò non basta a capire se fosse un bipede abituale o obbligato, sappiamo almeno che era capace di stare in piedi.
Il cranio, analizzato virtualmente, è stato "smontato" lungo le principali crepe, liberato dalle deformazioni tafonomiche e ricostruito per rappresentare la sua forma originaria. Si nota innanzitutto che il cranio nel suo complesso appare più largo, con un contorno occipitale del sagittale più arrotondato e un piano nucale orientato in modo più orizzontale. Le orbite sono più grandi e di forma più circolare, mentre la faccia è più alta nella direzione supero-inferiore. Mostra diverse caratteristiche derivate condivise con gli ominini successivi, come una faccia relativamente verticale con una premascella corta anteroposteriore, un forame magno posizionato anteriormente collegato a un basioccipitale relativamente corto e un piano nucale relativamente piatto, ampio e orientato orizzontalmente. Queste caratteristiche, insieme ad altre caratteristiche dentali come lo spessore dello smalto, supportano la conclusione che Sahelanthropus sia un ominino. L'ipotesi che Sahelanthropus fosse bipede, una caratteristica importante degli ominini, è un po' azzardata da fare perché sono le componenti post-craniali, come il femore, a fornire maggiori informazioni sulla locomozione. Tuttavia, ciò non esclude che il cranio possa fornire alcune informazioni biomeccaniche sulla specie in questione, ad esempio riguardo all'orientamento del collo. Esistono differenze sostanziali nell'orientamento del collo tra gli esseri umani e i primati non umani. Entrambi i gruppi riescono a muoversi con i loro piani orbitali, cioè attraverso la linea che unisce il margine inferiore e superiore delle orbite, approssimativamente perpendicolari al suolo. I primati, inoltre, orientano le vertebre cervicali superiori in modo più o meno perpendicolare al piano del forame magno, con un intervallo limitato di circa 10° di flessione ed estensione possibile a livello dell'articolazione cranio-cervicale. Nel caso degli esseri umani, l'angolo tra il forame magno e il piano orbitale è quasi perpendicolare (103,2 ± 6,9), mentre in P. troglodytes e in altre specie con posture pronograde, è di 63,7 ± 6,2°. In S. tchadensis, questo angolo è di circa 95° ed è simile a quello degli esseri umani e dei successivi ominini come A. afarensis ed A. africanus. Considerando che come quadrupede richiederebbe un angolo del collo molto esteso rispetto al piano del forame magno. Nonostante l'aumento di volume cerebrale rispetto alla lunghezza della base del cranio sia stato implicato nella rotazione orizzontale della base cranica posteriore, Sahelanthropus possedeva una capacità cranica di 360-370 cc, una delle più piccole nel lignaggio umano, risultando simile a quella dello scimpanzé. Tuttavia, il piano nucale piatto e orientato a circa 36° rispetto all'orizzontale di Francoforte lo avvicinano molto ad Homo e ad Australopithecus, allontanandola da Pan. Ciò, però, non fornisce molte informazioni sulla locomozione di questa specie.
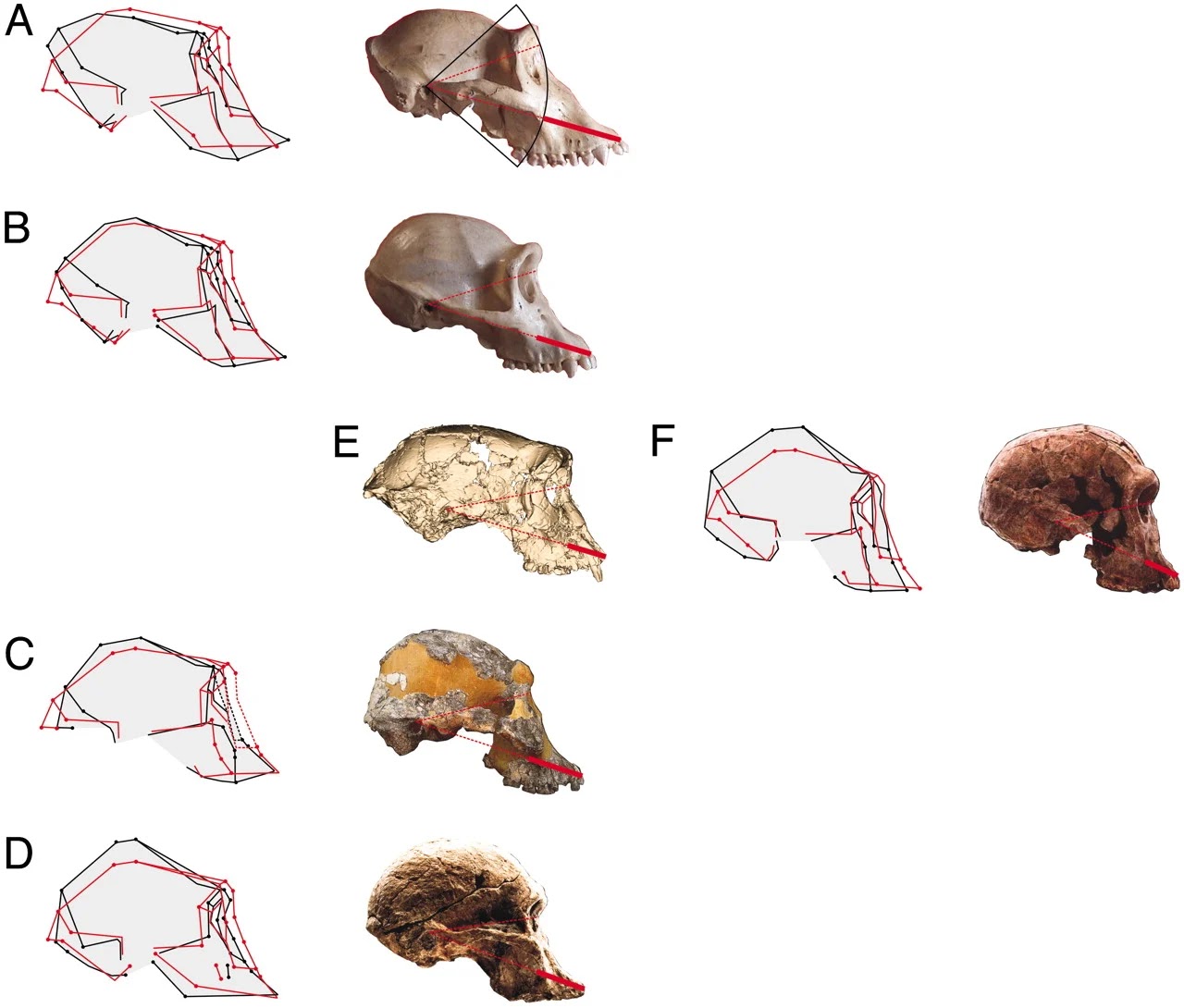 |
| Sovrapposizione GLS e dei crani lateralmente. Gorilla(A), Pan troglodytes(B), Australopithecus afarensis (C), Australopithecus africanus (D), Sahelanthropus tchadensis (E), Homo habilis (F), Guy et al,, 2005 |
 |
| Ricostruzione del cranio di Sahelanthropus tchadensis e quello non ricostruito confrontati con quelli di uomo e scimpanzé in orientamento orizzontale con la linea di Francoforte (Wolfpoff et al., 2006) |
Vediamo cosa dice lo studio del 2005 sui denti. Il corpo del frammento mandibolare di S. tchadensis è più gracile, con una larghezza massima del corpo a primo molare inferiore perpendicolare all'altezza del corpo di 14,5 mm, rispetto ad altri esemplari. Presenta un grande foro mentale situato nella parte centrale del corpo sotto il quarto premolare mandibolare. Il margine anteriore della sinfisi è verticale, con la parte inferiore inclinata posteriormente. Il piano alveolare della sinfisi è inclinato di circa 45° rispetto al piano alveolare del corpo. I tori trasversali inferiore e superiore sono debolmente sviluppati (quello superiore è più grande) e delimitano una fossa genioglossa poco profonda con un ampio foro genioglosso. La corona del canino è piccola con un contorno asimmetrico in vista occlusale a livello della cervice, rotta apicalmente e usurata distalmente. La corona del canino è corta, ma la sua radice è sorprendentemente lunga. Il quarto premolare mandibolare è subrettangolare, con un talonide ben sviluppato distolingualmente e un trigonide allungato, parzialmente conservato, con un metaconide distinto posizionato distalmente. Il terzo premolare mascellare presenta un contorno della corona occlusale ovale, con una leggera concavità sulla superficie mesiale sotto la cresta marginale. La superficie mesiale dello smalto mostra una faccetta di usura canina interprossimale ben delimitata sotto la cresta marginale mesiale, confermando l'assenza di un diastema tra il primo canino inferiore e il terzo premolare inferiore. Lo spessore dello smalto cuspidale postcanino in questi esemplari di S. tchadensis è intermedio tra i valori degli scimpanzé e le australopitecine.
 |
| Alcuni denti di Sahelanthropus tchadensis (Brunet et al., 2002) |
 |
| Porzione mandibolare con la serie dentale che parte dal terzo premolare al terzo molare. A destra la ricostruzione avvenuta attraverso la tomografia computerizzata. La barra misura 4 cm (Brunet et al., 2005) |
Sahelanthropus comprende ora un minimo di sei individui (un massimo di nove) provenienti da tre siti in una piccola area dell'unità di antracoteridi. S. tchadensis condivide importanti caratteristiche derivate con altri ominini, tanto da essere considerata come tale. La specie risulta essere vicina all'ultimo antenato comune degli scimpanzé e degli esseri umani. Nella dentatura, questi caratteri anatomici costituiscono un complesso canino-terzo premolare inferiore non levigato; assenza di diastema tra canino e terzo premolare inferiore; una sinfisi verticale con deboli tori trasversali; canini con corona piccola e radice lunga, corona canina inferiore con grosso tubercolo distale, entrambe le spalle molto basse; un terzo premolare superiore con superficie vestibolare fortemente inclinata; denti postcanini con spessore massimo dello smalto radiale intermedio tra scimpanzé e le australopitecine; e morfologia occlusale postcanina bulbosa e leggermente crenulata. Tutti gli esemplari di premolari mandibolari di ominino provenienti da Toros-Menalla hanno lo stesso modello di radice, con due radici e tre canali pulpari separati in ciascun premolare (uno mesiale e due distali) che mantengono la presunta condizione primitiva per il clade Pan-Homo. I caratteri anatomici del nuovo materiale di S. tchadensis, come una corona canina inferiore con una distinta cresta marginale mesiale e una striscia di usura scanalata distale che termina su un grande tubercolo distale, confermano le differenze morfologiche della specie del Ciad rispetto alle scimmie africane e le sue affinità morfologiche con il clade degli ominidi.
 |
| (a) vista frontale, (b) corona posteriore del canino sinistro (capovolta orizzontalmente), (c), Vista vestibolare sinistra. (d), Scansione mediosagittale della sinfisi (M, mesiale; D, distale). (e) vista occlusale. (f) Ricostruzione tridimensionale ; Vista disto-linguale del canino sinistro (g) e disegno (h) che mostra la posizione della striscia di usura distale e della rientranza. Barra della scala, 4 cm (Brunet et al., 2005) |
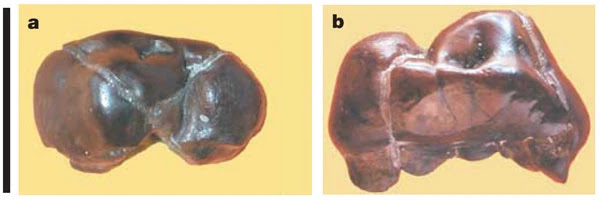 |
| Terzo premolare mascellare destro. (a) visione occlusale e (b) visione mesiale (Brunet et al., 2005) |
Bibliografia:
- Brunet, Guy, Pilbeam, Mackaye, Likius, Djimdoumalbaye, Beauvilain, Blondel, Bocherens, Boisserie, De Bonis, Coppens, Dejax, Denys, Duringer, Eisenmann, Gongdibé, Fronty, Geraads, Lehmann, Lihoreau, Louchart, Adoum, Merceron, Mouchelin, Otero, Pelaez Campomanes, Ponce De Leon, Rage, Sapanet, Schuster, Sudre, Tassy, Valentin, Vignaud, Viriot, Zazzo & Zollikofer (2002). A new hominid from the Upper Miocene of Chad, Central Africa, Nature, 418 (6894): 145-151.
- Brunet, M., Guy, F., Pilbeam, D. et al. New material of the earliest hominid from the Upper Miocene of Chad. Nature 434, 752–755 (2005).
- Daver, G., Guy, F., Mackaye, H.T. et al. Postcranial evidence of late Miocene hominin bipedalism in Chad. Nature 609, 94–100 (2022).
- Lebatard AE, Bourlès DL, Duringer P, Jolivet M, Braucher R, Carcaillet J, Schuster M, Arnaud N, Monié P, Lihoreau F, Likius A, Mackaye HT, Vignaud P, Brunet M. (2008) Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. Proc Natl Acad Sci U S A. 2008 Mar 4;105(9):3226-31.
- Michel Brunet, Alain Beauvilain, Denis Geraads, Franck Guy, Mahamat Kasser, Hassane T. Mackaye, Laura M. MacLatchy, Guy Mouchelin, Jean Sudre, Patrick Vignaud, Tchad: découverte d'une faune de mammifères du Pliocène inférieur, Comptes Rendus de l'Académie des Sciences - Series IIA - Earth and Planetary Science, Volume 326, Issue 2, 1998, Pages 153-158, ISSN 1251-8050
- N Patterson, DJ Richter, S Gnerre, ES Lander, D Reich, Genetic evidence for complex speciation of humans and chimpanzees. Nature 441, 1103–1108 (2006).
- Roberto Macchiarelli, Aude Bergeret-Medina, Damiano Marchi, Bernard Wood, Nature and relationships of Sahelanthropus tchadensis, Journal of Human Evolution, Volume 149, 2020, 102898, ISSN 0047-2484
- Suwa, G., Kono, R., Katoh, S. et al. A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448, 921–924 (2007).
- Vignaud, P., Duringer, P., Mackaye, H. et al. Geology and palaeontology of the Upper Miocene Toros-Menalla hominid locality, Chad. Nature 418, 152–155 (2002).
- Zollikofer C. P. E., Ponce de Leon M. S., Lieberman D. E., Guy F., Pilbeam D., Likius A., Mackaye H. T., Vignaud P. et Brunet M., 2005. Virtual reconstruction of Sahelanthropus tchadensis. Nature, 434 (7034): 755-759.



Nessun commento:
Posta un commento